量子生物學(xué)的未來:量子理論如何幫助理解生命?
點(diǎn)擊: 作者:Adriana Marais等 來源:人工智能學(xué)家 發(fā)布時(shí)間:2022-11-04 09:03:54

導(dǎo)語
2022年諾貝爾物理學(xué)獎(jiǎng)授予了關(guān)于量子信息科學(xué)的基礎(chǔ)性研究。一百多年前,量子革命為我們帶來了晶體管和激光,今天,基于量子信息的新技術(shù)正在讓我們進(jìn)入一個(gè)新的量子信息時(shí)代。事實(shí)上,已有研究表明,在生命過程中也存在量子現(xiàn)象。將量子理論應(yīng)用于生物學(xué)方面的研究被稱為“量子生物學(xué)”。
英國(guó)皇家學(xué)會(huì)會(huì)刊(Journal of the Royal Society Interface)在2018年11月發(fā)表綜述文章《量子生物學(xué)的未來》,給生物學(xué)家提供了一份簡(jiǎn)明的量子力學(xué)導(dǎo)論,介紹了量子生物學(xué)的幾個(gè)重要領(lǐng)域,包括光合作用、生物感知、酶催化反應(yīng)、大腦意識(shí)的涌現(xiàn)和生命起源等,提供了對(duì)量子生物學(xué)當(dāng)前發(fā)展?fàn)顩r的看法,并指出該領(lǐng)域要取得進(jìn)一步發(fā)展的潛在途徑。集智俱樂部對(duì)這篇文章做了全文翻譯。
研究領(lǐng)域:量子生物學(xué),光合作用,酶催化,感知,生命起源


目錄:
1. 簡(jiǎn)介
2. 什么是量子生物學(xué)?
3. 量子力學(xué):給生物學(xué)家的導(dǎo)論
4. 輸運(yùn)過程
5. 感知
6. 生命起源
7. 量子生物學(xué)和復(fù)雜性
生物系統(tǒng)是動(dòng)態(tài)的,不斷與環(huán)境交換能量和物質(zhì),以保持與生命同義的非平衡狀態(tài)。觀測(cè)技術(shù)的發(fā)展使研究人員能夠在越來越小的尺度上研究生物動(dòng)力學(xué)。這些研究在一系列生物過程中揭示了經(jīng)典物理學(xué)無法解釋的量子力學(xué)效應(yīng)的證據(jù)。量子生物學(xué)是對(duì)此類過程的研究,本文提供了該領(lǐng)域當(dāng)前狀態(tài)的概述,以及對(duì)未來方向的見解。
1. 簡(jiǎn)介
量子力學(xué)是描述亞原子粒子、原子、分子、分子組裝等微觀粒子屬性的基本理論。量子力學(xué)在納米和亞納米尺度上運(yùn)行,是光合作用、呼吸和視覺等基本生命過程的基礎(chǔ)。在量子力學(xué)中,所有物體都具有波動(dòng)特性:當(dāng)它們相互作用時(shí),量子相干性描述了由于波動(dòng)特性導(dǎo)致的描述這些物體的物理量之間的相關(guān)性。
在光合作用、呼吸和視覺方面,過去開發(fā)的模型基本上是量子力學(xué)的。研究人員已經(jīng)描述了基于表面跳躍框架的能量轉(zhuǎn)移和電子轉(zhuǎn)移。這些模型描述的動(dòng)力學(xué)通常是“指數(shù)”的,并且遵循費(fèi)米黃金法則[1,2]的應(yīng)用。由于是在準(zhǔn)連續(xù)的最終態(tài)的分布上求平均轉(zhuǎn)移率,計(jì)算的動(dòng)力學(xué)不再顯示相干性和干涉現(xiàn)象。在1990年代進(jìn)行的大量研究中,研究人員已經(jīng)在光合反應(yīng)中心和光捕獲復(fù)合體中觀察到了振蕩現(xiàn)象,通常將其歸因于振動(dòng)或混合電子-振動(dòng)波包的形成。據(jù)報(bào)道,在光合系統(tǒng)的激發(fā)能量轉(zhuǎn)移過程中檢測(cè)到非常長(zhǎng)壽命(660fs和更長(zhǎng))的電子量子相干性,重新激發(fā)了人們對(duì)“非平凡”量子力學(xué)在解釋生物體基本生命過程中的作用的興趣[3]。然而,量子現(xiàn)象(如相干性)可能在宏觀生命系統(tǒng)中發(fā)揮功能作用的想法并不新鮮。1932年,在量子物理學(xué)家尼爾斯·玻爾(Niels Bohr)因其在原子結(jié)構(gòu)方面的工作而獲得諾貝爾獎(jiǎng)10年后,他在哥本哈根國(guó)際光療法大會(huì)上發(fā)表了題為“光與生命”的演講[4]。這提出了一個(gè)問題,即量子理論是否有助于對(duì)生命系統(tǒng)的科學(xué)理解。出席會(huì)議的有一位充滿興趣的年輕物理學(xué)家 Max Delbrück,他后來幫助建立了分子生物學(xué)領(lǐng)域,并因在遺傳學(xué)方面的發(fā)現(xiàn)于1969年獲得諾貝爾獎(jiǎng)[5]。
所有生命系統(tǒng)都由分子組成,基本上所有分子都由量子力學(xué)描述。然而,傳統(tǒng)上,量子力學(xué)描述的系統(tǒng)與生物學(xué)研究的系統(tǒng)之間巨大的尺度差異,生命和無生命看似不同的性質(zhì),使得兩個(gè)知識(shí)體系之間一直存在某種隔閡。最近,超快光譜[6]、單分子光譜[7-11]、時(shí)間分辨顯微鏡[12-14]和單粒子成像[15-18]等實(shí)驗(yàn)技術(shù)的發(fā)展,使得研究人員能夠在越來越小的分辨率兼時(shí)間尺度上研究生物動(dòng)力學(xué)。他們揭示了生命系統(tǒng)功能所必需的各種過程,這些過程依賴于量子和經(jīng)典物理效應(yīng)之間微妙的相互作用。
量子生物學(xué)將量子理論應(yīng)用于經(jīng)典物理學(xué)無法準(zhǔn)確描述的生物學(xué)方面。盡管已有這個(gè)簡(jiǎn)單的定義,關(guān)于該領(lǐng)域在科學(xué)界的目標(biāo)和作用仍然存在爭(zhēng)議。本文提供了對(duì)量子生物學(xué)當(dāng)前地位的看法,并指出在該領(lǐng)域取得進(jìn)一步進(jìn)展的潛在途徑。
2. 什么是量子生物學(xué)?
就目前的范式而言,生物學(xué)在將經(jīng)典模型應(yīng)用于生命系統(tǒng)方面取得了廣泛的成功。在大多數(shù)情況下,分子尺度上的微妙量子效應(yīng)在整體生物功能中不發(fā)揮決定性作用。在這里,“功能”是一個(gè)廣義的概念。例如:視覺和光合作用如何在分子水平和超快時(shí)間尺度上發(fā)揮作用?由相距約0.3nm的堆疊的核苷酸分子組成的DNA,如何處理紫外線光子?酶如何催化重要的生化反應(yīng)?我們的大腦如何以亞納米尺度組織的神經(jīng)元處理如此驚人的信息量?DNA復(fù)制和表達(dá)是如何工作的?當(dāng)然,所有這些生物學(xué)功能都應(yīng)該在進(jìn)化適應(yīng)性的背景下考慮。在這些情況下,經(jīng)典近似和量子力學(xué)模型之間的差異通常被認(rèn)為可以忽略不計(jì),即使在基礎(chǔ)上,每個(gè)過程都完全受量子力學(xué)定律的支配。在量子和經(jīng)典疆域之間不明確的邊界上會(huì)發(fā)生什么?更重要的是,是否存在“看似”經(jīng)典但實(shí)際上并非如此的基本生物學(xué)功能?量子生物學(xué)的作用恰恰是揭露和解開這種聯(lián)系。
從根本上說,所有物質(zhì)——有生命或無生命的——都是量子力學(xué)的,由離子、原子和/或分子構(gòu)成,其平衡特性由量子理論精確確定。因此,可以說所有的生物學(xué)都是量子力學(xué)的。然而,這個(gè)定義并沒有解決生物過程的動(dòng)力學(xué)性質(zhì),或者分子間動(dòng)力學(xué)的經(jīng)典描述似乎往往已經(jīng)足夠這一事實(shí)。因此,量子生物學(xué)應(yīng)該根據(jù)所使用模型的物理“正確性”以及特定生物過程的經(jīng)典模型與量子力學(xué)模型的解釋能力的一致性來定義。
當(dāng)研究人員在納米尺度和更大尺度上研究生物系統(tǒng)時(shí),他們發(fā)現(xiàn)生物有機(jī)體中存在本文詳細(xì)介紹的過程。目前認(rèn)為,量子力學(xué)描述對(duì)于充分表征相關(guān)子系統(tǒng)的行為是必要的。雖然在宏觀時(shí)間和長(zhǎng)度尺度上很難觀察到量子效應(yīng),但總體上需要該過程。因此,生物體的功能和生存似乎依賴于分子間尺度的動(dòng)態(tài)量子力學(xué)效應(yīng)。量子生物學(xué)研究的正是這些時(shí)間和長(zhǎng)度尺度之間的相互作用,目的是建立一致的物理圖景。
對(duì)量子生物學(xué)的巨大希望可能包括對(duì)生命的定義和理解做出貢獻(xiàn),或者對(duì)大腦和意識(shí)的理解做出貢獻(xiàn)。然而,這些問題與科學(xué)本身一樣古老,更好的方法是詢問量子生物學(xué)是否可以為一個(gè)框架做出貢獻(xiàn),在該框架中研究人員可以以這樣的方式解決問題以獲得新的答案。對(duì)在量子物理學(xué)和經(jīng)典物理學(xué)領(lǐng)域之間有效運(yùn)行的生物過程的研究已經(jīng)有助于改進(jìn)對(duì)這種量子到經(jīng)典轉(zhuǎn)變的物理描述。
更直接的是,量子生物學(xué)有望為受生物啟發(fā)的量子納米技術(shù)提出設(shè)計(jì)原則,例如能夠在室溫下的嘈雜環(huán)境中,基于基本水平上有效執(zhí)行,甚至利用這些“嘈雜環(huán)境”來保護(hù)甚至增強(qiáng)量子特性[19,20]。通過設(shè)計(jì)這樣的系統(tǒng),有可能測(cè)試和量化量子效應(yīng)在多大程度上可以增強(qiáng)生物學(xué)中發(fā)現(xiàn)的過程和功能,并最終回答這些量子效應(yīng)是否可能是在系統(tǒng)設(shè)計(jì)中被有目的地選擇的。然而,重要的是,量子生物啟發(fā)技術(shù)也可以在本質(zhì)上獨(dú)立于啟發(fā)它們的生物體而有用。
3. 量子力學(xué):給生物學(xué)家的導(dǎo)論
二十世紀(jì)初,經(jīng)典物理學(xué)在描述所有可觀測(cè)現(xiàn)象方面的成功在某些方面開始受到挑戰(zhàn)。1900年,為了解釋黑體輻射的光譜能量分布,普朗克提出了一個(gè)想法,即物質(zhì)與頻率為 ν 的電磁輻射之間的相互作用是量子化的,僅以 hν 的整數(shù)倍發(fā)生,其中h是基本的普朗克常數(shù)。五年后,愛因斯坦進(jìn)一步發(fā)展了能量量子化的概念,將其擴(kuò)展到包括光子——光的量子。這個(gè)概念可以通過光電效應(yīng)來說明,其中入射到材料上的光會(huì)導(dǎo)致電子的發(fā)射。然而,即使是合適頻率的低強(qiáng)度光也會(huì)導(dǎo)致電子發(fā)射,而低于此閾值頻率的高強(qiáng)度光卻不起作用。為了解釋這個(gè)問題,愛因斯坦提出,在這種情況下光表現(xiàn)為粒子而非波的行為,離散能量可以轉(zhuǎn)移到材料中的電子。此外,玻爾在1913年提出的氫原子模型及其離散能態(tài),以及康普頓1923年對(duì)X射線的研究,都為現(xiàn)代物理學(xué)新時(shí)代的開始做出了貢獻(xiàn)。這些解釋黑體輻射和光電效應(yīng)以及原子穩(wěn)定性和光譜學(xué)的方法導(dǎo)致了量子力學(xué)的發(fā)展,這一理論在預(yù)測(cè)和描述微物理系統(tǒng)方面非常成功[21,22]。
普朗克和愛因斯坦假設(shè)輻射也表現(xiàn)出類似粒子的行為,從而開啟了量子革命,而德布羅意在1923年提出了補(bǔ)充理論——物質(zhì)本身具有波動(dòng)特性,波長(zhǎng)通過普朗克常數(shù)與其動(dòng)量相關(guān)。這個(gè)假說表明,物質(zhì)波應(yīng)該表現(xiàn)出衍射行為,隨后的實(shí)驗(yàn)證明了這一點(diǎn),表明電子等粒子顯示出干涉圖案。薛定諤在他的量子力學(xué)公式中建立了這一觀察結(jié)果,該公式通過使用波動(dòng)力學(xué)描述微觀系統(tǒng)的動(dòng)力學(xué)。量子力學(xué)公式允許研究量子態(tài)的許多重要方面:它在任何時(shí)間t的數(shù)學(xué)描述,如何計(jì)算與該狀態(tài)相關(guān)的不同物理量,以及如何描述狀態(tài)隨時(shí)間的演變[21,22]。
量子力學(xué)是一種數(shù)學(xué)框架,用完全刻畫系統(tǒng)的量子態(tài) |ψ> ,以及包含系統(tǒng)可用的所有狀態(tài)的希爾伯特空間 H,來描述一個(gè)孤立物理系統(tǒng)。由狀態(tài)向量 |ψ> 描述的系統(tǒng)的時(shí)間演化服從薛定諤方程,即
 (3.1)
(3.1)
其中 ? 是約化普朗克常數(shù),H(t) 是哈密頓量,代表系統(tǒng)的能級(jí)和系統(tǒng)各部分之間的相互作用。
量子理論更令人著迷的一個(gè)方面是,對(duì)于描述系統(tǒng)的兩個(gè)量子態(tài) |ψ1>和 |ψ2>,其線性疊加 α1 |ψ1> + α2 |ψ2> 也描述了該系統(tǒng)。構(gòu)成希爾伯特空間的狀態(tài)的這種組合或疊加更一般地寫為
 (3.2)
(3.2)
疊加態(tài)中的每個(gè)正交歸一態(tài) |ψi> 由復(fù)數(shù) αi 加權(quán),pi = |αi2> 是在該狀態(tài)找到系統(tǒng)的概率。正是在這里,量子力學(xué)與經(jīng)典世界分道揚(yáng)鑣。量子體系中疊加態(tài)的存在導(dǎo)致了獨(dú)特的量子力學(xué)性質(zhì),這通常是違反直覺的。例如,量子相干性描述了構(gòu)成疊加的各個(gè)態(tài)之間的明確關(guān)系,而量子糾纏是量子態(tài)之間一種特殊的關(guān)聯(lián)形式。一個(gè)很好的例子是光合作用中電子態(tài)的離域化,這對(duì)于解釋光合作用底層的電子能量轉(zhuǎn)移和電荷分離的速度和效率至關(guān)重要[23]。量子力學(xué)的概率詮釋也允許用隧穿的概念來預(yù)測(cè)量子粒子穿過勢(shì)壘的可能性,這在經(jīng)典力學(xué)中是被禁止的[21,22]。量子隧穿的一個(gè)很好的例子是供體和受體之間的電子有效轉(zhuǎn)移,如在光合作用反應(yīng)中心和呼吸復(fù)合物中,電子必須穿過高能蛋白屏障[24]。
在這篇綜述中,疊加、相干、糾纏和隧穿效應(yīng)這些特定的量子特性,被描述為對(duì)光合作用、磁感知、嗅覺、酶催化、呼吸和神經(jīng)傳遞等多種生物現(xiàn)象的新理解不可或缺的一部分[25-27]。
4. 輸運(yùn)過程
從根本上講,生物系統(tǒng)的動(dòng)力學(xué)與能量和電荷轉(zhuǎn)移有關(guān),后者涉及電子、質(zhì)子和離子。光合作用中的激發(fā)能量轉(zhuǎn)移和電荷轉(zhuǎn)移是量子生物學(xué)最成熟的領(lǐng)域,最新的研究領(lǐng)域是酶催化研究,它通常依賴電子和質(zhì)子的耦合來控制多個(gè)電荷的輸運(yùn)。
4.1.光合作用
超過30億年時(shí)間里,有一類多產(chǎn)的生物依靠光能生存,利用一種稱為光合作用的過程將光能轉(zhuǎn)化為對(duì)自身生存有用的形式。在典型條件下,光合生物將太陽光的能量轉(zhuǎn)化為生物質(zhì)的總體效率相當(dāng)?shù)?/span>(最多只有幾個(gè)百分點(diǎn)[28,29]),因?yàn)榧葻o法吸收所有入射的陽光,也無法將所有吸收的能量轉(zhuǎn)化為生物質(zhì)。然而,光合作用的初級(jí)光捕獲階段以近乎完美的量子效率進(jìn)行。這意味著在最佳條件下(低光強(qiáng)度和無壓力),對(duì)于幾乎每個(gè)被光捕獲(天線)復(fù)合物吸收和轉(zhuǎn)移的光子,都有一個(gè)電子在光合反應(yīng)中心內(nèi)轉(zhuǎn)移。
光捕獲復(fù)合物可能是準(zhǔn)無序的自組裝聚集體[30],或與蛋白質(zhì)支架結(jié)合的高度對(duì)稱的色素結(jié)構(gòu) [31,32]。構(gòu)成整個(gè)天線系統(tǒng)的光捕獲復(fù)合物與反應(yīng)中心相連,反應(yīng)中心是光合作用裝置中能夠進(jìn)行跨膜電荷分離的唯一單元[33]。
通過從自然光合系統(tǒng)中分離光捕獲復(fù)合物和反應(yīng)中心,同時(shí)模擬生理環(huán)境條件進(jìn)行的實(shí)驗(yàn),我們對(duì)光合光收集系統(tǒng)在體內(nèi)的功能有了更多了解。眾所周知,初級(jí)光合作用過程中的能量和電荷轉(zhuǎn)移表現(xiàn)出基本的量子力學(xué)特性,例如離域化、波包、相干性和超輻射,而且經(jīng)典模型不能準(zhǔn)確地描述隨后的動(dòng)態(tài)。
4.1.1.激發(fā)能量轉(zhuǎn)移
早在1938年,Franck&Teller[34]就提出光合作用中激發(fā)能量轉(zhuǎn)移的量子相干機(jī)制。他們考慮了Frenkel 激子的擴(kuò)散,這是單個(gè)光合色素電子激發(fā)的相干疊加(圖1)。隨著1990年代初期飛秒瞬態(tài)吸收光譜的出現(xiàn),細(xì)菌和植物光捕獲復(fù)合物中檢測(cè)到長(zhǎng)壽命振動(dòng)相干性,持續(xù)時(shí)間為皮秒級(jí)[35-40]。在過去14年中,先進(jìn)的超快技術(shù),稱為二維電子光譜(2D-ES),已被用于監(jiān)測(cè)光捕獲復(fù)合物振動(dòng)態(tài)和電子振動(dòng)(混合激子-振動(dòng))態(tài)的相干疊加的衰減。二維光譜顯示存在隨時(shí)間振蕩的交叉峰[41][42]。大量研究將交叉峰與激子態(tài)之間的耦合聯(lián)系起來,而它們的振蕩被分配給電子量子相干性,即激子態(tài)之間的相干疊加[42]。第一項(xiàng)此類研究是在綠色硫細(xì)菌的Fenna-Matthews-Olson(FMO)復(fù)合體上進(jìn)行的,本項(xiàng)研究由Fleming小組于2007年發(fā)表,揭示了特定的相干性持續(xù)時(shí)間極長(zhǎng)(660fs)[3]。2009年,高等植物的主要光捕獲復(fù)合物(LHCII)顯示出類似的振蕩信號(hào),并被解釋為量子相干能量轉(zhuǎn)移[43]。這些初始結(jié)果是在低溫下獲得的。一個(gè)重要的進(jìn)展是在2010年,Engel[44]和Scholes[45]小組獨(dú)立地檢測(cè)到FMO中生理溫度下類似的相干振蕩和兩種海洋隱藻藻類的光捕獲復(fù)合物。

圖 1. 展示光合作用光捕獲復(fù)合物中激發(fā)能量轉(zhuǎn)移概念的示意圖。橢圓描繪了其中發(fā)生強(qiáng)激子耦合的顏料簇,箭頭表示簇之間能量的不連貫轉(zhuǎn)移。在每個(gè)簇內(nèi),能量在所有顏料上都是離域的,并相干地傳遞。
通常,與低溫下的量子相干相比,生理溫度下的量子相干是很脆弱的,因?yàn)榄h(huán)境噪聲隨著溫度升高而增加,導(dǎo)致退相干時(shí)間更短。因此,這些結(jié)果很有趣,一方面從量子信息處理的角度來看,一個(gè)主要挑戰(zhàn)是在不可避免地與環(huán)境相互作用的系統(tǒng)中保持量子相干性,另一方面從量子生物學(xué)的角度來看,它研究生命系統(tǒng)功能的基本方面是否只能用量子力學(xué)來解釋。
受這個(gè)溫暖、嘈雜、復(fù)雜但非常有效的能量傳輸系統(tǒng)啟發(fā),人們提出了許多環(huán)境輔助量子輸運(yùn)模型。目的是將光合色素蛋白質(zhì)中多個(gè)發(fā)色團(tuán)位點(diǎn)的量子相干現(xiàn)象,與激發(fā)能量轉(zhuǎn)移過程(通常在系統(tǒng)的近似自旋玻色子模型內(nèi))的極端效率聯(lián)系起來。與噪音會(huì)降低系統(tǒng)性能的直覺相反,已經(jīng)發(fā)現(xiàn)與環(huán)境的相互作用可以提高傳輸效率[19,20]。光捕獲復(fù)合物由許多具有通常不同位點(diǎn)能量的色素分子組成。如果該能量差大于色素分子-色素分子耦合,則轉(zhuǎn)變將被抑制。移相噪聲可以改變位點(diǎn)能量,從而幫助克服這些能隙,并在不損失系統(tǒng)激發(fā)的情況下改善位點(diǎn)之間的輸運(yùn)[46]。
在支持和反對(duì)長(zhǎng)壽命(最多幾皮秒)電子相干性的大量理論證據(jù)之后,最近對(duì)FMO復(fù)合體的2D-ES研究明確表明,這些長(zhǎng)壽命的相干性主要來自基態(tài)振動(dòng)而不是激子–激子疊加[47,48]。發(fā)現(xiàn)與電子相干相關(guān)的量子拍頻幅度很小,衰減僅在60-240fs內(nèi)。然而,也發(fā)現(xiàn)了激發(fā)態(tài)振動(dòng)相干性的實(shí)驗(yàn)證實(shí)[48],其在光合作用中激發(fā)能量轉(zhuǎn)移中的作用仍有待確定。
然后人們可能會(huì)問,量子相干性——激子以及電子振動(dòng)相干性——在光收集中的作用是否是最優(yōu)化,或者僅僅是色素-蛋白質(zhì)復(fù)合物中色素接近的結(jié)果。還研究了環(huán)境輔助輸運(yùn)相對(duì)于系統(tǒng)或環(huán)境特征變化的穩(wěn)健性,結(jié)果表明這種穩(wěn)健性可用于高效量子輸運(yùn)系統(tǒng)的設(shè)計(jì)[49,50]。光合作用中時(shí)間尺度的收斂已被提出作為“量子金發(fā)姑娘效應(yīng)”的一個(gè)例子:自然選擇傾向于將量子系統(tǒng)驅(qū)動(dòng)到一個(gè)參數(shù)集,其中產(chǎn)生的量子相干度“恰到好處”地實(shí)現(xiàn)最大效率和最優(yōu)控制[51]。
一般來說,對(duì)振蕩動(dòng)力學(xué)的觀察不足以排除對(duì)相同行為的經(jīng)典描述,光合作用中環(huán)境輔助傳輸?shù)牧孔幽P鸵膊⒎菦]有爭(zhēng)議。然而,最近的工作[52]明確表明,環(huán)境振動(dòng)運(yùn)動(dòng)的非經(jīng)典特性可能有助于光合LHC在亞皮秒時(shí)間尺度和室溫下的激發(fā)能量轉(zhuǎn)移。這些想法應(yīng)該通過檢查光合LHC的突變變體是否會(huì)改變量子相干度、相干壽命以及能量轉(zhuǎn)移效率來進(jìn)行實(shí)驗(yàn)驗(yàn)證。
4.1.2.電荷轉(zhuǎn)移
光合作用中的電荷分離是自然界中最有效的現(xiàn)象之一,其量子效率接近百分之百。促成過程發(fā)生在不同的時(shí)間尺度上,從亞皮秒到毫秒,涉及無序和相干之間的相互作用,由電子振動(dòng)狀態(tài)(振動(dòng)和激子狀態(tài)的混合)介導(dǎo)。電荷分離是理解量子物理學(xué)在生物學(xué)中的作用的一個(gè)很好的候選者。由于電荷分離發(fā)生在微秒時(shí)間尺度上,量子效應(yīng)在這樣的宏觀水平上通常不是直接可見的。然而,電荷分離機(jī)制可以用一系列不同的過程來表示,早期步驟在超快時(shí)間尺度上并且只涉及幾個(gè)分子,而整體效率取決于每個(gè)步驟。

圖2. 根據(jù)[23]中的模型,高等植物光系統(tǒng)II反應(yīng)中心的量子相干電荷分離示意圖,沿著至少兩個(gè)現(xiàn)有途徑之一。左邊是諧波電位上的四個(gè)波包,每個(gè)波包對(duì)應(yīng)于電荷分離路徑上的不同狀態(tài)。對(duì)于每個(gè)狀態(tài),右側(cè)顯示了電荷云的關(guān)鍵顏料和大致位置和符號(hào)。水平線描繪了右側(cè)所示相應(yīng)狀態(tài)的指定波長(zhǎng)處的吸收峰,而波浪箭頭描繪了與指示振動(dòng)模式的共振相互作用。符和*分別代表電荷轉(zhuǎn)移特性和激子特性。這四種狀態(tài)的性質(zhì)是: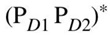 —高激子態(tài),
—高激子態(tài), —具有某些電荷轉(zhuǎn)移特性的激子態(tài),
—具有某些電荷轉(zhuǎn)移特性的激子態(tài), —與激子態(tài)混合的電荷轉(zhuǎn)移態(tài),
—與激子態(tài)混合的電荷轉(zhuǎn)移態(tài), ——最終的電荷分離狀態(tài)。請(qǐng)注意,最后兩個(gè)狀態(tài)之間涉及 ChlD1 的中間過程被省略了。由于與振動(dòng)模式耦合,前三個(gè)步驟涉及相干弛豫,而最后一步涉及非相干轉(zhuǎn)移。
——最終的電荷分離狀態(tài)。請(qǐng)注意,最后兩個(gè)狀態(tài)之間涉及 ChlD1 的中間過程被省略了。由于與振動(dòng)模式耦合,前三個(gè)步驟涉及相干弛豫,而最后一步涉及非相干轉(zhuǎn)移。
圖注:D1和D2,對(duì)稱反應(yīng)中心結(jié)構(gòu)的兩個(gè)分支;Chl,葉綠素;P,初級(jí)電子供體/特殊對(duì)Chl;Phe,脫鎂葉綠素。改編自Romero等人[23]。
1966年,DeVault和Chance觀察到紫色細(xì)菌反應(yīng)中心中電子轉(zhuǎn)移的溫度依賴性,這是經(jīng)典物理學(xué)無法解釋的[53]。他們提出這種行為作為量子隧穿的證據(jù)[54],并為生物學(xué)中電子和核隧穿的概念奠定了基礎(chǔ)[42]。雖然Marcus的電子轉(zhuǎn)移理論忽略了核隧穿[2],因此可能低估低溫下的電子轉(zhuǎn)移速率,但半經(jīng)典Marcus理論可以擴(kuò)展到基于非輻射躍遷理論的全量子力學(xué)處理,包括核隧穿,并且可以很好地預(yù)測(cè)電荷分離率隨溫度降低的增加。如果假設(shè)電荷分離與某些諧波振動(dòng)模式強(qiáng)耦合,則該速率由Jortner速率給出。然而,這是基于振動(dòng)弛豫發(fā)生在比電子轉(zhuǎn)移更短的時(shí)間尺度上的假設(shè),對(duì)于非常快速的轉(zhuǎn)移事件并非如此。超快光致電子轉(zhuǎn)移反應(yīng)如此之快,以至于在從供體到受體的電子轉(zhuǎn)移過程中不會(huì)發(fā)生完全的振動(dòng)弛豫。振動(dòng)弛豫通常發(fā)生在皮秒或亞皮秒時(shí)間尺度上,超快電子轉(zhuǎn)移反應(yīng)在相同尺度上進(jìn)行[55]。
高等植物光系統(tǒng)II反應(yīng)中心的2D-ES實(shí)驗(yàn)揭示了特定交叉峰的長(zhǎng)期振蕩,類似于光捕獲復(fù)合物,并被解釋為激子之間以及激子和電荷轉(zhuǎn)移之間的電子相干性狀態(tài)(圖2)[56]。在相干程度與有效和超快電荷分離之間觀察到很強(qiáng)的相關(guān)性[56]。實(shí)驗(yàn)結(jié)果由量子相干模型定量再現(xiàn),該模型具有新的能量轉(zhuǎn)移途徑,這些途徑在過多的振動(dòng)狀態(tài)之間通常不會(huì)出現(xiàn)[56-58]。具體來說,模擬表明觀察到的交叉峰振蕩可以通過特定的振動(dòng)模式維持,其中輔助共振轉(zhuǎn)移的模式主要是色素固有的,而色素激子躍遷主要由蛋白質(zhì)支架調(diào)節(jié)以匹配這些模式的能量。因此,雖然非平凡的量子效應(yīng)可能隱藏在宏觀層面,但它們似乎從根本上促進(jìn)了電荷分離的生物機(jī)制。因此,電荷轉(zhuǎn)移是理解量子物理學(xué)在生物學(xué)中作用的一個(gè)很好的過程。
因此,實(shí)驗(yàn)和理論證據(jù)表明,靜態(tài)無序景觀中的單一振動(dòng)模式很可能有助于光合生物中光收集和電荷分離的輸運(yùn),蛋白質(zhì)支架調(diào)節(jié)的色素激子躍遷與振動(dòng)模式的能量相匹配,有助于共振轉(zhuǎn)移[52,56]。核電子(振動(dòng))耦合是分子系統(tǒng)光誘導(dǎo)功能的重要機(jī)制,一般而言,在特定設(shè)計(jì)的系統(tǒng)中。光誘導(dǎo)電子轉(zhuǎn)移的結(jié)果也已被證明可以通過模式特定的紅外激發(fā)與電子轉(zhuǎn)移途徑耦合的振動(dòng)而發(fā)生根本性的改變[59]。
4.1.3.單分子光譜
單分子實(shí)驗(yàn)提供了一種有趣且有前景的方法來研究光合復(fù)合物的量子特征。2D-ES的結(jié)果表明,環(huán)境以分子振動(dòng)的形式對(duì)量子相干能量和電子轉(zhuǎn)移過程具有密切的作用。這種環(huán)境從復(fù)雜到復(fù)雜以及在相對(duì)較慢的時(shí)間尺度(毫秒到分鐘)上變化,正如對(duì)單個(gè)光捕獲復(fù)合物的研究中報(bào)道的不同的、隨時(shí)間變化的能量轉(zhuǎn)移途徑所表明的那樣[11, 60-62]。由于這種異質(zhì)性,通常稱為“靜態(tài)紊亂”,起源于相對(duì)緩慢的蛋白質(zhì)構(gòu)象動(dòng)態(tài)波動(dòng),光譜特征振蕩的衰減受到系綜平均的強(qiáng)烈影響。通過避免單分子實(shí)驗(yàn)中的系綜平均,可以將量子相干性擴(kuò)展到微觀領(lǐng)域,這可能會(huì)更多地揭示這種量子現(xiàn)象的生理意義。
靜態(tài)無序還通過改變激發(fā)被困在能量匯中的概率[63,64],或不與光系統(tǒng)中其他復(fù)合物相鄰的色素簇[65]來強(qiáng)烈影響光捕獲復(fù)合物中激發(fā)的命運(yùn)。對(duì)光捕獲復(fù)合物的外部影響對(duì)光動(dòng)力學(xué)的精細(xì)控制仍缺乏詳細(xì)了解,例如在非光化學(xué)猝滅期間發(fā)生的光動(dòng)力學(xué),這是一種耗散光合作用中多余能量的重要調(diào)節(jié)機(jī)制,但目前仍然缺乏了解。振動(dòng)輔助能量轉(zhuǎn)移和能量轉(zhuǎn)移途徑采樣的作用可以通過單分子相干控制實(shí)驗(yàn)來闡明。
最近,基于對(duì)光合作用反應(yīng)中心磁場(chǎng)效應(yīng)的一系列實(shí)驗(yàn)觀察,自旋的量子力學(xué)性質(zhì)在光合作用的量子保護(hù)機(jī)制中的直接作用已被提出[66]。了解監(jiān)管和保護(hù)光合作用的機(jī)制,以及這些過程到飛秒級(jí)時(shí)間尺度上的分辨率和控制,對(duì)于在干旱脅迫或高太陽輻射條件下的作物歉收調(diào)查,以及下一代仿生太陽能電池的開發(fā)中具有重要應(yīng)用。
獲取有關(guān)單個(gè)分子中純量子力學(xué)特征信息的一種補(bǔ)充方式是尋找不同的量子力學(xué)指紋,例如高階相關(guān)函數(shù)中的非經(jīng)典亞泊松統(tǒng)計(jì),或某些違反貝爾不等式或柯西-施瓦茨不等式的可觀測(cè)量的統(tǒng)計(jì)。最初熱化振動(dòng)的非經(jīng)典性已被證明是通過相干激子-振動(dòng)相互作用引起的,并且明確地由與電子動(dòng)力學(xué)耦合的有效集體模式的相空間準(zhǔn)概率分布,即使激發(fā)的非相干輸入也是如此。這些結(jié)果表明,對(duì)輔助激發(fā)和電荷傳輸、光接收和化學(xué)傳感過程的振動(dòng)運(yùn)動(dòng)的非經(jīng)典特性的研究,可能是揭示生物學(xué)中非平凡量子現(xiàn)象的作用的試金石[52,67]。然而,由于對(duì)超快(飛秒到納秒)時(shí)間分辨率和高光子計(jì)數(shù)率的要求,在單分子水平上進(jìn)行此類研究極具挑戰(zhàn)性。
4.1.4.人工光合作用
量子生物學(xué)領(lǐng)域的一項(xiàng)主張是,在微觀尺度上詳細(xì)了解光合作用,尤其是其初級(jí)階段,將使研究人員能夠設(shè)計(jì)受生物啟發(fā)的人工光合作用系統(tǒng),利用地球上豐富的元素(如碳)來有效利用太陽光、氧氣、氮?dú)獾取@此類系統(tǒng)的設(shè)計(jì)已經(jīng)取得了重大進(jìn)展(例如[68-73])。人工光合作用的分子方法是將太陽能轉(zhuǎn)化為生物燃料的眾多競(jìng)爭(zhēng)者之一。在這里,研究自然光合作用以提取設(shè)計(jì)原理,然后嘗試基于這些原理開發(fā)更好的光系統(tǒng)。當(dāng)前系統(tǒng)的主要限制是存儲(chǔ)以及由過度積累電荷引起的催化劑的不穩(wěn)定性。電荷分離的平衡是必不可少的,例如通過耦合質(zhì)子運(yùn)動(dòng)[74]。
受量子生物學(xué)啟發(fā)的設(shè)計(jì)中的一個(gè)重要原則是激子的存在:它們負(fù)責(zé)更有效的光吸收、更快的能量漏斗(即沿能量梯度衰減更快)、更快的能量轉(zhuǎn)移和更有效(不可逆)的激發(fā)捕獲反應(yīng)中心[75]。此外,它們的量子行為通過它們對(duì)不同路徑的波動(dòng)干涉,以及蛋白質(zhì)環(huán)境的振動(dòng)模式與色素相互作用的方式(增加離域和提取或吸收振動(dòng)能量),以促進(jìn)能量在復(fù)合物中的轉(zhuǎn)移。
對(duì)原型人工反應(yīng)中心的初級(jí)電荷轉(zhuǎn)移的研究揭示出,電子和原子核在幾十飛秒的時(shí)間尺度上的關(guān)聯(lián)波動(dòng)作為光致電流產(chǎn)生循環(huán)的驅(qū)動(dòng)機(jī)制[76]。重要的是在評(píng)估此類系統(tǒng)時(shí)包括周轉(zhuǎn)率,而不僅僅是效率。良好的周轉(zhuǎn)率需要有彈性、穩(wěn)定的系統(tǒng),這可能可以通過某種保護(hù)性聚合物包裹顏料來實(shí)現(xiàn)[77]。例如,已經(jīng)研究了制造涉及跨膜BChl-結(jié)合蛋白模型的人工膜嵌入蛋白的策略[78]。
光合作用中激子傳輸研究的挑戰(zhàn),包括色素網(wǎng)絡(luò)的無序性質(zhì)、類似時(shí)間尺度上激子能級(jí)的波動(dòng)以及與激子動(dòng)力學(xué)的耦合,也出現(xiàn)在其他非生物環(huán)境中。證明最初出現(xiàn)在量子生物學(xué)中的見解和想法很有用的一個(gè)具體例子是最近對(duì)有機(jī)光伏(PV)的研究,這是一種太陽能電池,由兩種不同的人造分子半導(dǎo)體組合而成。電子供體體半導(dǎo)體通常是聚合物,而受體通常是富勒烯衍生物[6,6]-苯基-C61-丁酸甲酯。在器件內(nèi)部產(chǎn)生強(qiáng)束縛的 Frenkel 激子,然后擴(kuò)散直到它們到達(dá)供體和受體區(qū)域之間的界面。供體和受體具有不同的化學(xué)勢(shì),這提供了將界面處的激子解離成自由電荷的驅(qū)動(dòng)力,然后自由電荷可以擴(kuò)散到相反的電極。然而,由于庫(kù)侖相互作用是長(zhǎng)程的,并且有機(jī)材料具有低介電常數(shù),電子和空穴在室溫下經(jīng)歷比kBT高10倍的結(jié)合能,即使它們已經(jīng)離解到界面兩側(cè)的不同分子上。因此,絕大多數(shù)設(shè)備從電極中提取的電荷遠(yuǎn)少于入射光產(chǎn)生的電荷,從而導(dǎo)致功率轉(zhuǎn)換效率低下。
然而,近年來出現(xiàn)了少數(shù)高效器件,據(jù)報(bào)道電荷提取效率接近100%[79,80]。研究人員已經(jīng)進(jìn)行了大量工作來確定該子集中有效的遠(yuǎn)程電荷分離的起源。實(shí)驗(yàn)工作表明,有機(jī)光伏和光合作用有一些驚人的巧合。最初認(rèn)為供體-受體界面處的電荷分離將發(fā)生在皮秒到納秒的時(shí)間尺度上,這可以通過經(jīng)典的熱漲落驅(qū)動(dòng)的電荷跳躍來描述。許多實(shí)驗(yàn)已經(jīng)觀察到,在激子到達(dá)界面的100飛秒內(nèi)產(chǎn)生了很大一部分的自由電荷,就像在光合激子傳輸過程中觀察到的飛秒相干性和動(dòng)力學(xué)一樣。在經(jīng)典的Marcus“跳躍”模型中無法解釋這種時(shí)間尺度上的傳輸,必須明確包含相干傳輸[81,82]。
理解這些觀察結(jié)果需要對(duì)供體-受體界面附近的電荷輸運(yùn)進(jìn)行詳細(xì)建模。至關(guān)重要的是,有機(jī)半導(dǎo)體中的電荷移動(dòng)經(jīng)歷了上述色素-蛋白質(zhì)復(fù)合物的所有三個(gè)關(guān)鍵特性:高度無序、分子波動(dòng)和重組。因此,也許是出現(xiàn)類似的現(xiàn)象并不奇怪。光合作用中的激子傳輸理論通常涉及部分離域狀態(tài)和噪聲輔助傳輸?shù)南嗷プ饔茫枋隽巳绻ぷ踊螂姾膳c分子振動(dòng)模式適度強(qiáng)耦合,以及如何增強(qiáng)通過網(wǎng)絡(luò)的傳輸。越來越多的共識(shí)認(rèn)為,部分離域狀態(tài)也出現(xiàn)在高效的有機(jī)光伏系統(tǒng)中,并且也有人認(rèn)為受主半導(dǎo)體聚集區(qū)域內(nèi)的噪聲使電荷分離過程成為可能。許多研究有機(jī)光伏的理論小組現(xiàn)在采用開放量子系統(tǒng)方法,使用的方法受到早期光合光捕獲復(fù)合物研究的強(qiáng)烈影響[83-86]。
量子生物學(xué)有助于開發(fā)可持續(xù)能源技術(shù)的潛力通常被認(rèn)為是研究該領(lǐng)域的動(dòng)力,特別是用于理解光合作用系統(tǒng)中的量子相干激發(fā)能量轉(zhuǎn)移。然而,從這一普遍主張到實(shí)用能源系統(tǒng)的發(fā)展,需要更具體的建議,將量子能量轉(zhuǎn)移到環(huán)境可持續(xù)和經(jīng)濟(jì)可擴(kuò)展的系統(tǒng)中。正如量子生物學(xué)本身的一個(gè)基本問題一樣,納米尺度現(xiàn)象如何能夠顯著影響生物體的宏觀行為,在工程宏觀能源系統(tǒng)中利用量子生物現(xiàn)象的潛力取決于這種現(xiàn)象是否能在環(huán)境和經(jīng)濟(jì)約束下顯著影響系統(tǒng)規(guī)模的運(yùn)行。硅基光伏等現(xiàn)有可再生能源技術(shù)的發(fā)展表明,能源轉(zhuǎn)換效率等純物理性能標(biāo)準(zhǔn)通常不如生命周期分析等技術(shù)經(jīng)濟(jì)性能指標(biāo)重要。現(xiàn)有的極其高效的光伏電池對(duì)于大多數(shù)實(shí)際應(yīng)用來說過于昂貴,并且在某些情況下需要低豐度的元素,即使負(fù)擔(dān)得起,它們也無法在全球范圍內(nèi)擴(kuò)展。解決這些限制一直是最近光伏研究的核心問題,其他全球范圍的考慮對(duì)于評(píng)估量子生物學(xué)在可持續(xù)能源發(fā)展中的潛力很重要。在全球范圍內(nèi),只有不到20%的能源作為電力被消耗;幾乎所有剩余的80%都在最終消耗時(shí)作為燃料消耗。盡管作為電力消耗的能源份額穩(wěn)步增長(zhǎng),但在人為氣候變化和化石燃料枯竭的臨界點(diǎn)的時(shí)間范圍內(nèi),燃料可能仍然占主導(dǎo)地位。因此,迫切需要可再生燃料技術(shù),例如工程和/或人工光合作用系統(tǒng),而不是簡(jiǎn)單的更高效的光伏電池發(fā)電。開發(fā)光合能源系統(tǒng)需要一種集成的系統(tǒng)方法,光收集組件只是復(fù)雜(生物)物理化學(xué)系統(tǒng)的一部分,理想情況下應(yīng)該針對(duì)系統(tǒng)規(guī)模性能進(jìn)行優(yōu)化。只有通過平衡子系統(tǒng)的還原分析與整個(gè)系統(tǒng)的整體分析,才能公平地評(píng)估量子動(dòng)力學(xué)效應(yīng)在此類系統(tǒng)內(nèi)的光收集中的重要性。這樣的考慮有助于集中主要?jiǎng)訖C(jī)是可持續(xù)能源發(fā)展的量子生物學(xué)研究人員的努力。關(guān)鍵的教訓(xùn)是,為了將全球系統(tǒng)規(guī)模的目標(biāo)與能源系統(tǒng)(包括在量子動(dòng)力學(xué)現(xiàn)象起著重要作用的納米級(jí))范圍內(nèi)的工程參數(shù)相關(guān)聯(lián),集成系統(tǒng)方法至關(guān)重要。
4.2.酶催化
酶通過催化生化反應(yīng)對(duì)細(xì)胞功能至關(guān)重要,否則這些生化反應(yīng)的反應(yīng)速率可能非常低。理解速率加速的物理機(jī)制是一個(gè)困難的研究課題。過渡態(tài)理論已被用作解釋酶催化的基礎(chǔ),但最近的理論和實(shí)驗(yàn)發(fā)展開始關(guān)注量子隧穿效應(yīng)在酶催化多氫轉(zhuǎn)移中的潛在作用,并且一直專注在量子力學(xué)形式中描述氫轉(zhuǎn)移[87]。描述量子隧穿的標(biāo)準(zhǔn)模型已被證明可以解釋實(shí)驗(yàn)酶數(shù)據(jù),只要能解釋酶具有許多可能的不同結(jié)構(gòu)的事實(shí)[88]。有趣和熱門的問題包括,蛋白質(zhì)的局部振動(dòng)運(yùn)動(dòng)是否可以通過與反應(yīng)坐標(biāo)偶聯(lián)來為酶提供催化優(yōu)勢(shì),以及是否可以選擇蛋白質(zhì)的特定動(dòng)力學(xué)運(yùn)動(dòng)來輔助催化。
酶通常依靠電子和質(zhì)子的耦合來控制電荷傳輸和催化作用[87]。在生物能量?jī)?chǔ)存中,由電子轉(zhuǎn)移驅(qū)動(dòng)的質(zhì)子易位的重要性于1961年首次被注意到[89],從那時(shí)起,質(zhì)子耦合電子轉(zhuǎn)移機(jī)制[90]已被證明是氨基酸自由基生成和運(yùn)輸?shù)幕A(chǔ)[91],伴隨著酶活性位點(diǎn)的大多數(shù)底物鍵的激活[92]。
5. 感知
生命系統(tǒng)會(huì)根據(jù)從環(huán)境感知中獲得的信息不斷更新內(nèi)部過程。環(huán)境中的微小變化都可能導(dǎo)致生物體功能發(fā)生宏觀變化。有人提議說,幾種生物傳感機(jī)制非常敏感,可以在量子水平上檢測(cè)環(huán)境中的變化。下面的小節(jié)概述了這些建議。
5.1.磁感知
鳥類遷徙的量子理論并不是一個(gè)新理論。自Schulten首次提出以來40年[93],自由基對(duì)機(jī)制在鳥類磁感知的實(shí)驗(yàn)和理論研究中都得到了很好的確立,并且是用作量子生物學(xué)領(lǐng)域證據(jù)的主要替代(光合作用)例子之一:該機(jī)制可分為三個(gè)步驟(圖3)。在光激發(fā)導(dǎo)致電子轉(zhuǎn)移和對(duì)形成的初始步驟之后,自由基對(duì)在單態(tài)和三重態(tài)自旋狀態(tài)之間振蕩,然后再進(jìn)行重組和神經(jīng)解釋的最后一步。正是單態(tài)-三重態(tài)混合經(jīng)歷了地球磁場(chǎng)的影響,并提供了一些解釋行為觀察的方法,例如光依賴[94]、傾斜方面[95]和鳥類羅盤的共振效應(yīng)[96,97]。
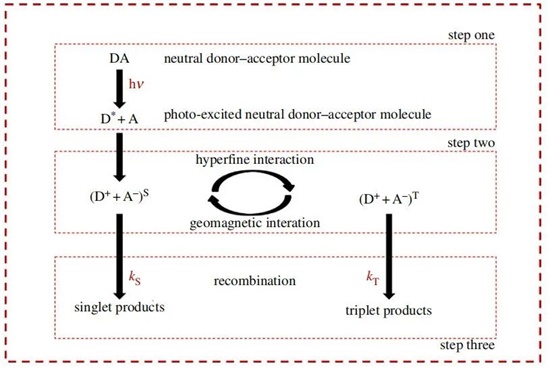
圖3.自由基對(duì)機(jī)制的三個(gè)步驟。在第一步中,入射到供體分子上的光子導(dǎo)致電子對(duì)中的一個(gè)電子被提供給受體,以產(chǎn)生空間分離但自旋相關(guān)的自由基對(duì)。在第二步中,自由基對(duì)在塞曼效應(yīng)和超精細(xì)效應(yīng)的影響下在單態(tài)和三重態(tài)之間振蕩。第三步包括化學(xué)產(chǎn)品中的自旋相關(guān)的重組。
自由基對(duì)理論已經(jīng)產(chǎn)生了許多關(guān)于羅盤相干性和糾纏的論文,并為羅盤行為提供了令人信服的論據(jù)(參見[98]和其中的參考文獻(xiàn)以了解該主題)。然而,對(duì)于羅盤機(jī)制是否真的是一種量子現(xiàn)象或可以使用半經(jīng)典框架來描述,仍然存在一些疑問。對(duì)此有人提出,指南針令人具有難以置信的準(zhǔn)確性,是由于避免了隱花色素中形成的自由基的自旋能級(jí)交叉,這是一個(gè)真正的量子力學(xué)過程[99]。
在解決機(jī)制結(jié)構(gòu)方面也取得了進(jìn)展。隱花色素作為磁感受位點(diǎn)的生物分子,目前受到廣泛關(guān)注。有證據(jù)表明,在植物中,弱磁場(chǎng)會(huì)增強(qiáng)隱花色素的反應(yīng)[100]。在果蠅的案例中,也曾有研究人員提出隱花色素可能介導(dǎo)動(dòng)物的磁反應(yīng)[101,102]。對(duì)于更復(fù)雜的生物來說,這似乎是合乎邏輯的下一步,特別是因?yàn)橐呀?jīng)在候鳥眼中確認(rèn)了四種不同類型的隱花色素[103]。并且,已經(jīng)證明來自候鳥的隱花色素在藍(lán)色光譜的影響下,會(huì)形成具有毫秒壽命的自由基[104]。最近,兩項(xiàng)研究為這四種隱花色素之一Cry4在鳥類羅盤中發(fā)揮作用提供了強(qiáng)有力的證據(jù)。Cry4以恒定水平而非周期性模式表達(dá);這種恒定性對(duì)于高效導(dǎo)航是必要的[105]。Cry4的雙錐定位,以及它在候鳥遷徙期間表達(dá)水平上調(diào)但在雞中不上調(diào)的事實(shí)進(jìn)一步證實(shí)了該分子作為磁感受器是可行的[106]。
鳥類遷徙作為量子生物學(xué)主題的未來將依賴于對(duì)地球磁場(chǎng)中隱花色素的自旋動(dòng)力學(xué)的更詳細(xì)了解,以及這如何解釋已經(jīng)提出的一些行為問題,其中之一是如何調(diào)和對(duì)應(yīng)用振蕩射頻場(chǎng)的迷失方向影響的不同觀察結(jié)果。除其他外,隱花色素負(fù)責(zé)調(diào)節(jié)晝夜節(jié)律,與DNA修復(fù)酶光解酶非常相似[107,108]。更好地了解隱花色素與地球磁場(chǎng)的相互作用,可以進(jìn)一步了解更復(fù)雜的生物體中可能表現(xiàn)出量子效應(yīng)的其他生物過程。
5.2.嗅覺
嗅覺是生物體“聞”數(shù)千種不同分子的系統(tǒng)。與人類味覺結(jié)合的數(shù)百種不同類型受體和果蠅中的數(shù)十種受體如何產(chǎn)生如此驚人靈敏的分子識(shí)別系統(tǒng)仍然是一個(gè)謎。1928年首次提出的一個(gè)有趣的理論[109]是,我們的嗅覺依賴于氣味分子的量子力學(xué)振動(dòng)模式。1996年,這一理論再次復(fù)興,它提出蛋白質(zhì)偶聯(lián)受體使用非彈性電子隧穿來測(cè)量分子振動(dòng),而不是分子和受體像鎖和鑰匙一樣只是形狀匹配 [110]。根據(jù)這一理論,只有當(dāng)這些位點(diǎn)之間的能量差與氣味劑的振動(dòng)能量匹配時(shí),電子才會(huì)從供體位點(diǎn)隧穿傳輸?shù)绞荏w位點(diǎn)。這種嗅覺振動(dòng)理論讓人想起光合作用中激子的聲子輔助傳輸,說明量子化振動(dòng)在量子生物學(xué)中的基本作用。

圖4.說明量子效應(yīng)在嗅覺中的可能作用的示意圖。先前已提出嗅覺取決于適合特定受體的氣味劑的形狀和大小,而果蠅實(shí)驗(yàn)表明,盡管氘和氫兩種同位素具有相同的形狀,但用氘取代氫會(huì)導(dǎo)致氣味發(fā)生變化。這表明振動(dòng)頻率可能在氣味檢測(cè)中發(fā)揮作用[111]。
為支持該理論,果蠅實(shí)驗(yàn)表明氣味劑的形狀和大小不足以檢測(cè)。例如,盡管氘和氫兩種同位素分子具有相同的形狀(圖4)[111],但用氘取代氫會(huì)導(dǎo)致氣味發(fā)生變化。此外,已經(jīng)觀察到具有相似振動(dòng)頻率的分子會(huì)引起果蠅的相似反應(yīng),盡管它們?cè)诨瘜W(xué)上無關(guān)[111]。2007年[112]顯示了使用簡(jiǎn)單但通用的理論模型提出的機(jī)制的物理可行性和效率。下一步是設(shè)計(jì)受生物學(xué)啟發(fā)的室溫分子傳感設(shè)備。
5.3.認(rèn)知
量子物理學(xué)是否可以在解決尚未解決的身心問題中發(fā)揮作用的問題,即大腦的形態(tài)結(jié)構(gòu)如何產(chǎn)生了有意識(shí)的思考,這絕不是新問題。Michael Lockwood 1989年的思想、身體和量子[113]或Henry Stapp 于2009年Mindful Universe中的觀點(diǎn)[114]是一些具有哲學(xué)或科普角度的專著的例子。大量的關(guān)注和批評(píng)也指向羅杰·彭羅斯和 Stuart Hameroff 的想法,他們提出神經(jīng)細(xì)胞中細(xì)胞骨架的一部分,即微管,根據(jù)他們所謂的“精心策劃的客觀減少”(orchestrated objective reduction)進(jìn)行量子計(jì)算,這是一種來自彭羅斯的量子引力機(jī)制[115]。
另一個(gè)科學(xué)分支從更實(shí)用的角度觸及“量子大腦”的問題:量子神經(jīng)網(wǎng)絡(luò)研究試圖利用量子計(jì)算來改進(jìn)廣泛用于機(jī)器學(xué)習(xí)的人工神經(jīng)網(wǎng)絡(luò)模型。這些人工神經(jīng)網(wǎng)絡(luò)模型歷史上源自生物神經(jīng)網(wǎng)絡(luò)的動(dòng)力學(xué)[116,117],因此接近量子動(dòng)力學(xué)和神經(jīng)計(jì)算的兼容性問題。然而,MaxTegmark[118]提出了一個(gè)關(guān)鍵點(diǎn),他估計(jì)參與動(dòng)作電位傳播的離子的退相干時(shí)間尺度比神經(jīng)動(dòng)力學(xué)的相關(guān)時(shí)間尺度小10~20個(gè)數(shù)量級(jí)。換句話說,參與神經(jīng)動(dòng)力學(xué)的離子的量子相干性會(huì)在宏觀動(dòng)力學(xué)之前很久就被破壞了可能會(huì)受到影響。因此,生物神經(jīng)網(wǎng)絡(luò)中量子效應(yīng)的潛在理論必須展示生物神經(jīng)網(wǎng)絡(luò)的宏觀動(dòng)力學(xué)如何從小得多的相干動(dòng)力學(xué)中產(chǎn)生。Matthew Fisher 在這個(gè)方向上進(jìn)行了有希望的研究,他提出磷可以充當(dāng)神經(jīng)量子比特,允許在大腦中進(jìn)行量子處理,并且這種量子性受到所謂的波斯納分子的保護(hù),波斯納分子將磷酸根離子與鈣離子結(jié)合。糾纏的波斯納分子然后觸發(fā)神經(jīng)元放電率的非局部量子相關(guān)性[119]。
麻醉藥的研究引導(dǎo)了研究意識(shí)的實(shí)驗(yàn)之路。到目前為止,關(guān)于意識(shí),研究人員只能說它“可溶于氯仿”[120],并且可以溶于一系列彼此截然不同的分子中。在最近一篇關(guān)于意識(shí)研究的文章中[121],Turin 和他的同事發(fā)現(xiàn),許多全身麻醉劑可逆地增加果蠅中的電子自旋含量,而這種作用在抗麻醉的突變果蠅中是不存在的。他們提出,全身麻醉劑會(huì)改變某些分子的最高占據(jù)分子軌道的結(jié)構(gòu),促進(jìn)供體和受體之間的電子傳輸,從而使大腦處于無意識(shí)狀態(tài)。如果這是真的,那么由密度泛函理論(量子理論的一種工具)計(jì)算出的分子結(jié)構(gòu)變化將對(duì)宏觀尺度產(chǎn)生影響。此外,電子自旋測(cè)量對(duì)于觀察這種效應(yīng)至關(guān)重要。尤其是在量子生物學(xué)的更廣泛定義中,此類發(fā)現(xiàn)非常令人鼓舞,并顯示了該學(xué)科可以做出的潛在貢獻(xiàn)。
6. 生命起源
原始生物(如細(xì)菌)中量子效應(yīng)的識(shí)別導(dǎo)致開放量子系統(tǒng)模型成功應(yīng)用于光合作用系統(tǒng)中的能量和電荷轉(zhuǎn)移過程,并表明量子效應(yīng)可能在從無生命物質(zhì)到由無生命物質(zhì)組成的最早的生命系統(tǒng)的涌現(xiàn)中發(fā)揮了重要作用。
對(duì)星際冰中生命分子前體的探測(cè)表明,生命的組成部分可能已經(jīng)出現(xiàn)在太空中,并由彗星或隕石等物體運(yùn)送到地球。然而,標(biāo)準(zhǔn)的計(jì)算量子化學(xué)無法解釋星際介質(zhì)中化學(xué)物質(zhì)的多樣性和豐富性。例如,氰化氫(HCN)低聚物可能在前生命分子(prebiotic molecule)的合成中發(fā)揮了重要作用[122-126],最近,在星際空間中檢測(cè)到了HCN的二聚體形式(氰基甲亞胺)[127]。然而,已經(jīng)發(fā)現(xiàn)星際氣相生產(chǎn)路線無法生產(chǎn)大量的HCN [128,129],盡管它可以被檢測(cè)到。
星際介質(zhì)中前生命分子自發(fā)產(chǎn)生的理論研究根本上是在開放量子系統(tǒng)的框架下進(jìn)行的。低溫星際冰冷環(huán)境與簡(jiǎn)單的分子系統(tǒng)如HCN強(qiáng)耦合,并被入射的紫外線輻射激發(fā),可以使用來自量子生物學(xué)領(lǐng)域的適當(dāng)?shù)谋娝苤慕浦颠M(jìn)行研究。
一個(gè)更困難的問題是,第一個(gè)生命系統(tǒng)是如何從這些前生命分子中產(chǎn)生的,如果有的話。盡管在可行的DNA設(shè)計(jì)[130]方面取得了非凡的工作,我們還不能從基本成分合成一個(gè)小的功能肽,并且對(duì)理解很多問題有很長(zhǎng)的路要走,比如如何準(zhǔn)確區(qū)分分子集合和組成生命系統(tǒng)的分子集合,以及量子效應(yīng)在生命起源中可能發(fā)揮了什么作用。
7. 量子生物學(xué)和復(fù)雜性
復(fù)雜性可以定義為系統(tǒng)描述的不可壓縮性。如果沒有可以簡(jiǎn)化問題的基本集合,則系統(tǒng)是復(fù)雜的。在宏觀生物系統(tǒng)中可見的量子特征必須能夠在過渡到高光譜密度的過程中幸存下來。頻譜中的混沌漲落包含有關(guān)系統(tǒng)動(dòng)力學(xué)的普遍特征的信息。量子生物學(xué)的新興領(lǐng)域關(guān)注長(zhǎng)度和時(shí)間尺度分離的動(dòng)態(tài)現(xiàn)象之間的相互作用,從納米尺度分子組裝中的飛秒能量轉(zhuǎn)移過程,到整個(gè)生物體尺度的生態(tài)系統(tǒng)內(nèi)的生存和繁殖。
重要的是要注意,自然選擇不能違反化學(xué)的量子力學(xué)性質(zhì)。例如,不可能設(shè)計(jì)出具有光捕獲復(fù)合物的植物,組成復(fù)合物的化合物卻不符合量子力學(xué)描述。生物學(xué)描述了在整個(gè)生物體的尺度上進(jìn)化選擇的系統(tǒng)。選擇的結(jié)果是通過遺傳過程介導(dǎo)的,對(duì)生物子系統(tǒng)的影響只能達(dá)到一個(gè)特定尺度,在該尺度之下發(fā)生的物理細(xì)節(jié)不受生物選擇的影響。然后,量子生物學(xué)關(guān)注可以選擇的量子動(dòng)力學(xué)過程(例如,在蛋白質(zhì)尺度上)是否會(huì)影響宏觀有機(jī)體動(dòng)力學(xué)。也就是說,如果納米尺度的量子行為要傳達(dá)選擇優(yōu)勢(shì),它必須是可選擇的,而且這種選擇發(fā)生在整個(gè)有機(jī)體的尺度上。因此,我們可能會(huì)聲稱,只有可以存在于量子和經(jīng)典變體中的子系統(tǒng)才是有意義的,因?yàn)檫M(jìn)化可以選擇其中之一。這使以下主張變得微不足道,即所有生物學(xué)都是量子生物學(xué),因?yàn)樗蕾囉诨瘜W(xué),而所有化學(xué)在原子和分子尺度上都是量子的;因此,進(jìn)化在這里不可能選擇量子子系統(tǒng)以外的任何東西。量子生物學(xué)感興趣的子系統(tǒng)可以居住在什么樣的長(zhǎng)度和時(shí)間尺度范圍內(nèi)(因?yàn)榭梢韵胂笏鼈兛赡艽嬖谟诹孔雍徒?jīng)典變體中)是一個(gè)懸而未決的問題。尺度問題將證明對(duì)量子生物學(xué)的進(jìn)一步發(fā)展很重要[131]。
8. 討論與結(jié)論
關(guān)于量子生物學(xué)的第一本書是 Pascual Jordan [132] 撰寫的《有機(jī)分子之謎的物理學(xué)》(Physics of the mystery of organic molecules)。然而,自1932年出版以來,關(guān)于生命本質(zhì)的許多謎團(tuán)仍然存在。很明顯,粗粒化的經(jīng)典模型無法準(zhǔn)確描述生命系統(tǒng)中發(fā)生的一系列過程。那么,持續(xù)爭(zhēng)論的問題是量子效應(yīng)在這種生物過程中發(fā)揮重要作用的程度。
回答這個(gè)問題的一個(gè)有用途徑是通過生物啟發(fā)量子技術(shù)的工程設(shè)計(jì),這些技術(shù)可以勝過為相同目的(例如用于能量利用或環(huán)境傳感)而設(shè)計(jì)的經(jīng)典設(shè)備。一些自然過程在生理溫度和極其復(fù)雜的系統(tǒng)中經(jīng)過了數(shù)十億年完善,如果宏觀尺度上的量子效應(yīng)可以在其中發(fā)揮作用,使其良好運(yùn)轉(zhuǎn),那么生物世界中存在大量信息可以從中為人類自己的技術(shù)汲取靈感。
在這個(gè)方向上,人們提出了一種原型量子熱機(jī),它清楚地說明了量子設(shè)計(jì)原理,即電子自由度和振動(dòng)自由度之間的單一能量量子的相干交換可以增強(qiáng)光收集系統(tǒng)的功率,超過單獨(dú)使用熱機(jī)制可能實(shí)現(xiàn)的功率。它使用熱力學(xué)性能測(cè)量的量子優(yōu)勢(shì)已被量化,并且該原理對(duì)現(xiàn)實(shí)生物結(jié)構(gòu)的適用性得到了證明[133]。
量子生物學(xué)研究生物功能和該功能的調(diào)節(jié),其與靜態(tài)無序相關(guān)。單分子光譜給我們一個(gè)獨(dú)特強(qiáng)大的透鏡來觀察靜態(tài)無序的作用,它連接生物功能(投影到宏觀/有機(jī)尺度)和量子力學(xué)現(xiàn)象。同時(shí),量子生物學(xué)還關(guān)注長(zhǎng)度和時(shí)間尺度分離的動(dòng)力學(xué)現(xiàn)象之間的相互作用,從納米級(jí)分子組裝中的飛秒能量轉(zhuǎn)移過程,到整個(gè)生物尺度內(nèi)生態(tài)系統(tǒng)中的生存和繁殖。
雖然在未來幾十年內(nèi),量子生物學(xué)將證明受生物啟發(fā)的量子設(shè)備在多大程度上優(yōu)于經(jīng)典類似物,但更深層次的問題是,納米尺度的量子動(dòng)力學(xué)現(xiàn)象如何為整個(gè)生物體提供選擇優(yōu)勢(shì)。嚴(yán)格解決這個(gè)問題需要說明,對(duì)有機(jī)體適應(yīng)性具有重要意義的宏觀物理可觀測(cè)量如何可預(yù)測(cè)地依賴于納米級(jí)量子動(dòng)力學(xué)變量。相反,我們還必須考慮,納米級(jí)的量子子系統(tǒng)如何通過進(jìn)化依賴于生物體的宏觀動(dòng)態(tài)。這個(gè)問題的進(jìn)展可能會(huì)得到一個(gè)理論框架的幫助,該框架允許有機(jī)體尺度模型通過納米尺度模型進(jìn)行參數(shù)化。這可以由復(fù)雜系統(tǒng)理論領(lǐng)域內(nèi)的多尺度分析工具提供。我們還可以設(shè)想一些實(shí)驗(yàn),在這些實(shí)驗(yàn)中,已知在納米尺度上表現(xiàn)出長(zhǎng)壽命量子相干過程的野生型生物與已知不存在此類過程的轉(zhuǎn)基因生物競(jìng)爭(zhēng)。這樣的實(shí)驗(yàn)——類似于生物學(xué)家定期進(jìn)行的那些實(shí)驗(yàn)——可以提供清楚的洞見,表明量子生物學(xué)現(xiàn)象是否可以為生物體提供選擇優(yōu)勢(shì),并增加量子生物學(xué)作為生物學(xué)的一個(gè)領(lǐng)域的可信度。
來源:集智俱樂部
作者:Adriana Marais, et al.
譯者:趙雨亭
責(zé)任編輯:向太陽
特別申明:
1、本文只代表作者個(gè)人觀點(diǎn),不代表本站觀點(diǎn),僅供大家學(xué)習(xí)參考;
2、本站屬于非營(yíng)利性網(wǎng)站,如涉及版權(quán)和名譽(yù)問題,請(qǐng)及時(shí)與本站聯(lián)系,我們將及時(shí)做相應(yīng)處理;
3、歡迎各位網(wǎng)友光臨閱覽,文明上網(wǎng),依法守規(guī),IP可查。
作者 相關(guān)信息
內(nèi)容 相關(guān)信息
? 昆侖專題 ?
? 高端精神 ?

? 新征程 新任務(wù) 新前景 ?


? 國(guó)策建言 ?

? 國(guó)資國(guó)企改革 ?

? 雄安新區(qū)建設(shè) ?

? 黨要管黨 從嚴(yán)治黨 ?

熱點(diǎn)排行
 剛剛,中國(guó)憋出大招!比美國(guó)傳統(tǒng)芯片快1000倍,外媒:拜登頭疼了
剛剛,中國(guó)憋出大招!比美國(guó)傳統(tǒng)芯片快1000倍,外媒:拜登頭疼了
 鐵礦石商全部投共,天地改換!
鐵礦石商全部投共,天地改換!
建言點(diǎn)贊
 高洪義等:著力發(fā)揮山東長(zhǎng)山列島在媽祖文化傳播中的新時(shí)代效應(yīng)
高洪義等:著力發(fā)揮山東長(zhǎng)山列島在媽祖文化傳播中的新時(shí)代效應(yīng) 秦安:股市暴跌!三大原因,兩個(gè)跡象,說明我們必須要打好金融戰(zhàn)
秦安:股市暴跌!三大原因,兩個(gè)跡象,說明我們必須要打好金融戰(zhàn) 習(xí)五一:互聯(lián)網(wǎng)管理規(guī)則應(yīng)確立“以人民為中心”的理念
習(xí)五一:互聯(lián)網(wǎng)管理規(guī)則應(yīng)確立“以人民為中心”的理念? 社會(huì)調(diào)查 ?
圖片新聞